X-inactivation
(XCI)
- X-inactivation
(XCI) occurs in somatic cells early in development (2nd week)
- Blastocyst stage:
- Stage
of development when embryo has < 100 cells
- XCI
was discovered by Lyon in 1961
- The
choice in which X is inactivated is apparently random
- All
subsequent cells arising from that cell will have the same X inactivated
- Females
are normally mosaic with respect to which X is inactivated
- Mechanism
of randomization is unclear
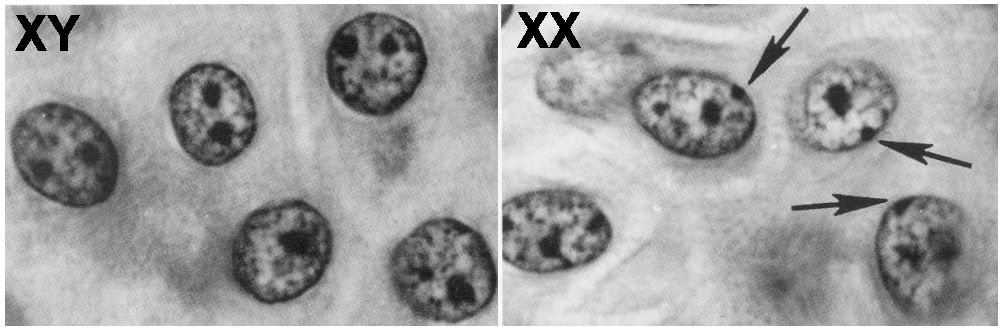
- The
inactivated X chromosome forms a heterochromatic mass (Barr body) in interphase cells
- Replicates
late during the cell cycle
- See
picture at: http://www.mun.ca/biology/scarr/Barr_Bodies.jpg
- Measuring
X-inactivation status:
- Replication
banding
- Only
a hundred or so cells can realistically be studied by this technique
- Molecular
methodology
- Large
numbers of cells are analyzed
- Androgen
receptor locus at Xq13 (close to the XIC) is often used
- Only the
tissue sampled is being tested actually – the other tissues may have
different inactivation patterns
- mechanism
of inactivation (see fig. 3 in Wutz & Gribnau, 2007)
- pathways
for gene silencing may be regulated in a cell type-specific manner
- X
inactivation centre (XIC) is mapped to proximal Xq
(see figure in Wutz 2007 or Tolland 2008)
- Xq13
- Regulates
3 basic steps of X-inactivation:
- counting
of the number of X chromosomes
- marking
one X chromosome to remain active
- Inactivating
the other X chromosome(s)
- Contains
XIST gene (Xi-specific
transcripts)
- Key
master regulatory locus for X inactivation
- X-inactivation
cannot occur in its absence
- Triggers
chromosome-wide gene repression
- Not
required for maintenance of X inactivation once it is established
- Only
expressed by allele on inactive X chromosome (Xi)
- Mbd2 is required for DNA
methylation to repress expression on the
active X (Xa)
- Cis-acting
- Acts
only on the chromosome it is actually on
- Product
is a noncoding RNA that stays in close
approximation with inactive X chromosome
- Xist forms a repressive
compartment from which the transcription machinery is exluded in early X inactivation
- LINE
repeats on X have been implicated in spreading inactivation across
the chromosome
- May
act as “way-stations” or “boosters”
- L1
LINEs are enriched on X chromosomes (nearly doubled)
- In
X:A translocations, XCI is incomplete and often discontinuous
- autosomes with fewer L1s
tend to be spared of inactivation
- high
densities of L1 are also associated with autosomal
mono-allelically expressed genes
- Possible
mechanism:
- L1s
could form secondary structures
- facilitation
of Xist spreading
- “capturing”
X-linked genes by physically consigning them to the silent
internal compartment
-
- TSIX
gene controls XIST expression in early stages of XCI
- Long
untranslated RNA
- Functions
to block X inactivation
- Negatively
regulates Xist
- Mechanism
of action of Tsix
on Xist
is unknown
- Transcribed
in antisense over XIST (see fig. in Wutz,
2007)
- Hypothesis
– Tsix blocks Xist function via an
RNA interference (RNAi) mechanism
- Ogawa
et al (2008) report Xist and Tsix duplexes in
vivo in mice
- Hypothesis
– the process of transcription of TSIX prevents the transcription of
XIST
- Homozygous
deletion of TSIX:
- Results
in equal proportions of cells in which one or both X chromosomes are
silenced
- Requires
the intronic DXPas34 element to function
- Minisatellite
- Deletion
of DXPas34 element results in nonrandom XCI of the mutated X
chromosome
- And
a defect in imprinted XCI
- Role
of Tsix-directed
chromatin change
- Recruitment
of specific regulatory factors and chromatin modifications to the
XIST promoter
- Tsix is in turn controlled by XITE locus
- Produces
several non-coding RNAs
- Contains
several strong DNase I hypersensitive sites
(DHS)
- XITE
enhancer enables Tsix to persist allele-specifically on the future Xa
- Tsix/Xite
pairing may be required to reliably block silencing on Xa
- Xce (X-controlling
element)
- Involved
in choice mechanism
- Transient
pairing of homologous XICs
occurs in female cells
- Occurs
after X-inactivation has been initiated but prior to the onset of
random XCI
- occurs
prior to Xist
accumulation and prior to establishment of epigenetic modifications
- Tsix and Xite are necessary and sufficient
for pairing
- XIC
pairs are localized close to the nuclear envelope
- Changes
in 3D organization of sequences due to pairing may impact gene
expression
- In
the extraembryonic tissues, the paternal X is
always silenced (imprinted XCI)
- Does
this impact imprinting assays on CVS’s?
- Hypotheses
for mechanism of choosing X for inactivation (see figure in Wutz, 2007):
- autosomal blocking factor (BF)
- protects
one X chromosome per diploid genome
- potential
blocking factor binding site – DXPas34
- located
at 3’ of XIST
- deletion
may result in ectopic XCI in male cells
- nature
of the BF remains elusive
- symmetry
breaking
- alternate
states
- cohesion
of sister chromatids is regulated
differently between the two female X chromosomes
- transvection
- both
XIC’s meet and decide the
future Xa and Xi
- XIC-XIC pairing has been observed
- But
XCI was not affected in female cells with a 65kb deletion 3’ of XIST which shows loss of XIC pairing
- Stochastic
- Each
X has a probability of being inactivated
- Methylation, acetylation,
and chromatin folding changes:
- In
addition to these genetic controls, a regulatory ciruit
dependent on primary chromosome structure has been described
- Various
temporal phases of XCI have been linked to a dynamic reorganization of
Xi internal structure
- Early
in X-inactivation
- Tsix activation affects
the level of histone H3 Lys methylation
- Alters
the chromatin structure of the XIST/TSIX locus
- Xist recruits polycomb group complexes (PRC1 and PRC2) to the Xi
- establishes
chromosome-wide histone modifications
- histone macroH2A is highly
enriched in inactive X chromatin
- thought
to create a repressive environment for gene expression
- Xi
is organized into an outer rim of genes and a repeat-rich core or
compartment
- Xist localizes to the
repeat-rich core
- genes
localize to the outside of this compartment
- genes
are recruited into this compartment as they are silenced (in general)
- genes
escaping X inactivation remain at the periphery of the Xi territory
- genes
are silenced in a manner that requires the Xist repeat-A sequence
- Exact
mode of action of Xist is unknown
- Unknown
how silencing is restricted to one specific chromosome
- promoter
region of many genes on the inactive X chromosome is modified by CpG island methylation
- enzyme:
DNA methyltransferase
- A
variety of chromatin modifiers (polycomb
group complexes, histone deacetylases,
and DNA methylases) maintain repression of
the Xi
- SAF-A
(attachment factor A)
- binds
satellite DNA and SARs/MARs (scaffold attachment regions / matrix
attachment regions)
- enriched
on Xi
- Mouse
model of X inactivation:
- Murine embryonic stem (ES)
cells
- Undifferentiated
ES cells:
- Low
level Xist
and Tsix
expression
- Upon
differentiation of ES cells:
- Tsix expression is
extinguished on the future Xi
- Xist expression is upregulated on the future Xa
(Xi?)
- not
all genes on X are subject to inactivation (see figure in Brown 2003)
- primary
pseudoautosomal region (PAR) (Xp22.3) (Y____)
- pseudoautosomal genes present on both
X and Y
- terminal
2.6 Mb of Xp
- homologous
with terminal Yp
- secondary
pseudoatosomal region
- 320
kb of terminal Xq
- Homologous
with terminal Yq
- at
least 15% of genes are expressed from both copies of X
- another
10% show variable X inactivation between individuals
- some
females inactivate them, some do not
- genes
that escape inactivation are expressed at a lower and level than their Xa
counterparts
- level
of expression is also variable
- location
of genes escaping inactivation is not random
- occur
in clusters
- local
chromatin structural changes likely responsible for clustering
- terminal
Xp is enriched for genes escaping
inactivation
- up
to 50% of genes, compared to a few percent on Xq
- ?
related to distance from XIST
- therefore,
imbalance for genes on Xp may have greater
clinical significance than imbalance involving Xq
- Inactivation
rate of genes appears to be related to evolution of X and Y chromosomes
- Genes
more recently lost from Y chromosome are more likely to escape
X-inactivation
- Genes
remotely lost from Y are less
likely to escape X-inactivation
- X-inactivation
in the presence of chromosomal imbalance and other genetic disorders:
- Show
calico cat
- X-linked
diseases:
- In
some disorders, females are mosaic
- Ex.
Duchenne muscular dystrophy
- Phenotypes
vary according to the proportion of cells with a mutant allele on the
active X in the relevant tissue
- Therefore,
dominance and recessiveness for X-linked
disorders is not absolute (variable penetrance)
- ~40%
of X-linked disorders are classified as recessive because they show
little or no penetrance in female heterozygotes
- Examples
- Hemophilia
A (factor VIII deficiency)
- X-linked
colour-blindness
- In
some disorders, there is skewed inactivation of the mutant allele
(in relevant tissues)
- Likely
due to survival disadvantage of cells expressing the mutant allele
- Can
be useful in diagnosis of carrier state of some X-linked disorders:
- X-linked
immunodeficiencies
- Dyskeratosis congenital
- Incontinentia pigmenti
- Manifesting
heterozygote:
- In
some disorders, “skewed” X inactivation can occur by chance
- The
fraction of cells in various tissues of carrier females in which
the normal or mutant allele happens to remain active can be quite
variable
- If
this skewed inactivation is present in the pertinent tissues, it can
cause a female carrier to manifest the disorder
- Degree
of penetrance varies from disorder to
disorder
- In
fragile X syndrome, nearly 50% of females show developmental
abnormalities
- Generally
to a lesser extent than males
- ~30%
are classified as dominant because they are penetrant
in most (>85%) female heterozygotes
- Phenotype
is usually milder in females
- The
mutant allele is located on the inactive X chromosome in a
proportion of cells
- Examples:
- X-linked
hypophosphatemic rickets
- ~30%
are penetrant in some (15-85%)
- X-linked
disorders with male lethality:
- X
chromosome abnormalities
- In
almost all patients with unbalanced structural abnormalities of an X
chromosome, the structurally abnormal chromosome is always the inactive
X
- Likely
reflects secondary selection against genetically unbalanced cells
- Accounts
for the increased frequency observed for unbalanced X abnormalities
- Aneuploidy:
- Only
one X per diploid genome is active, regardless of the number of X’s
- t(X;Autosome)
References:
·
Nussbaum RL, McInnes RR, Willard HF.
Thompson & Thompson Genetics in Medicine. 7th ed. Saunders; 2007.
·
Wutz A, Gribnau J. X inactivation Xplained.
Curr. Opin. Genet. Dev.
2007;17(5):387-93.
·
Tsai C, Rowntree RK, Cohen DE, Lee JT.
Higher order chromatin structure at the X-inactivation center via looping DNA. Dev. Biol. 2008;319(2):416-25.
·
Lyon MF. Gene action in the X-chromosome of the mouse (Mus musculus L.). Nature. 1961;190:372-3.
·
Carrel L, Willard HF. X-inactivation profile reveals extensive
variability in X-linked gene expression in females. Nature. 2005;434(7031):400-4.
·
Brown CJ, Greally JM. A stain upon the
silence: genes escaping X inactivation. Trends
Genet. 2003;19(8):432-8.
·
Abrams L, Cotter PD. Prenatal diagnosis of de novo X;autosome translocations. Clin. Genet. 2004;65(5):423-8.
·
Heard E. Recent advances in X-chromosome inactivation. Curr. Opin. Cell Biol.
2004;16(3):247-55.
·
Avner P, Heard E.
X-chromosome inactivation: counting, choice and initiation. Nat. Rev. Genet. 2001;2(1):59-67.
·
{kind=link}