Note: This Page was written as an introduction to the Page on Retinitis pigmentosa. If you are familiar with the anatomy of the eye and the normal functioning of the visual pathway please go to: Retinitis pigmentosa
The
Anatomy of the Eye
The
Fundus of the Eye
The
Cell Layers of the Retina
The
Photoreceptor Cell Layer
The
Proteins of the Outer Segments
Rhodopsin
The
Phototransduction Cascade
The Role of cGMP
in Vision
Metabolism
of Vitamin A in the Retina
Acknowledgements
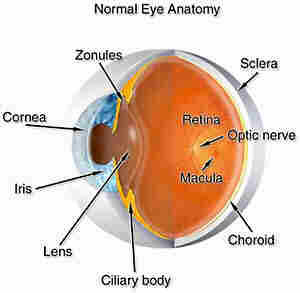
Figure 1. Shows a cross-section of an eyeball where the various anatomical structures of the eye are depicted.
Looking at a cross-section of the eyeball we see the following:
Damage to the stroma, Descemets membrane or the endothelium would cause permanent damage to the cornea. This damage would interfere with the transmission of light to the retina and thus, lead to blindness.
The cornea acts like a window which allows light to pass through several tissues in the eye before it arrives at the retina.
If cataract develops in the lens, light will not pass through to the retina and vision is blocked.
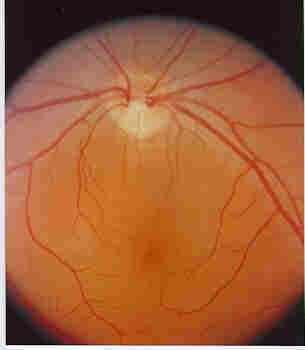
Figure 2.- The fundus of a normal Eye. Note the uniform color of the fundus, and the blood vessels which traverse the fundus starting at the pale colored Optic Disc seen at the top of the picture. The Optic Disc is the area from which the Optic nerve leaves the retina to connects with the brain. (Original slide provided by Dr. Gilles Desroches).
Looking into the eye with an ophthalmoscope, one sees the retina or what is referred to as the fundus. The retina surrounds the cavity of the eyeball. A number of blood vessels come out (the arteries) and leave (the veins) through a pale area in the retina, called the Optic Disc.
The Optic Nerve passes through the Optic Disc to transmit nerve impulses to the brain.
The Macula , is located in the fundus. Inside the macula is the Fovea where most of our sharp vision is located because of the concentration of visual cells, called the Cones, in that area (see later).
Pathological changes to the macula lead to the disease called Macular Degeneration.
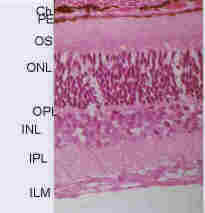
Figure 3. - The Cell Layers of the retina. Abbreviations: CH = choroid, PE = pigment epithelium, OS = outer segment layer, ONL = outer nuclear layer, OPL = outer plexiform layer, INL = inner nuclear layer, IPL = inner plexiform layer, ILM = inner limiting membrane.
If we were to look at a small thin specially treated and stained section of the wall of the eyeball, under the microscope, we see the following, starting from the outside of the eye (see Figure 3):
The photoreceptor cell layer can be divided into four sections:
Light is reflected from the object we are viewing, goes through the cornea and the lens where it is focused onto the retina.
At the retina, light passes through the inner retina to the photoreceptor cells where it is absorbed by the visual pigment, Rhodopsin, found in the outer segments of the photoreceptor cells.
Rhodopsin then undergoes a series of chemical reactions with other proteins found in the outer segments. These interactions,activate a number of photoreceptor outer segment enzymes, whose activities lead to the "hyperpolarization" of the photoreceptor cell.
The hyperpolarization of the visual cell cause the transmission of an electrical impulse which travels through the inner retina to the brain forinterpretation.
The photoreceptor cells consist of two types of cells:

Figure 4: Diagram of a cone cell showing outer segment with comb-like discs, nuclear areas and synaptic body. |
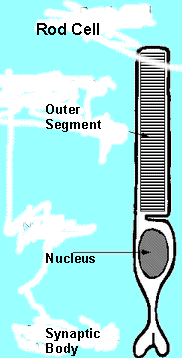
Figure 5 - Diagram of a rod cell showing the outer segment with its stack of discs separated from the cell membrane, the inner segment, the nucleus and the synaptic body. |
In spite of their importance, these cells are small in number in the mammalian retina and are mainly concentrated in the macula and particularly in the fovea.
Both the cone and the rod cells, are made up of four distinct areas (see Figures 4 and 5):
The cone and rod cells not only differ in their function but they also differ in the structure and shape of their outer segments. Thus:
The rod outer segmets, on the other hand are:

Figure 6 - An electron micrograph of a section of a rod outer segment. Note the double membranes of the discs and the fact that the discs are separated from the cell membrane.
The proteins of the outer photoreceptor segments are synthesized in the inner segments and exported by the Golgi Body to the outer segments. In the cones, the new proteins are layed in the outer segment area closest to the inner segments. In the rod cell the exported proteins are formed into discs which are layed at the bottom of the outer segment.
As new proteins are delivered to the outer segments the older ones are pushed towards the apex of the outer segments. Eventually, the oldest proteins are shed and are removed that is, phagocytised by the neighboring pigment epithelial cells (RPE), thus keeping the area between the tips of the outer segments and the RPE clean. In the RPE the phagocytised discs are digested and moved to the back of the RPE cell.
The process of phagocytosis occurs in a 24 hour circadian rhythm. In Rod cells it takes place daily at light onset and at sunset for the cones.
The pigment epithelium appears to have other importantfunctions. Thus, under intense light conditions, it extends long processes around the photoreceptor outer segments presumably to protect the outer segments from light damage. Under dim light conditions These processes are retracted to allow for more light to reach the outer segments.
RPE also plays a role in the process of vision whereby it is involved in the conversion of all-trans retinal to 11-cis retinal (see below)
The undulating membranes of the cones contains the photopigment, Photopsin, while the discs of the rods contain the visual pigment, Rhodopsin. Chemically, Photopsin and Rhodopsin seem to have little differences between them and often, they are referred to as Rhodopsin.
Rhodopsin forms about 80 to 90% of the proteins of the rod outer segment disks. It consists of a protein, called opsin, and a special form of vitamin A, called 11- cis retinal.
Opsin consists of a single polypeptide chain of 348 amino acids that winds itself seven times around the outer segment disc membrane forming four loops facing the outside of the ROS disc and three loops facing the inside of the disc.
The rhodopsin molecule is folded in a three dimensional manner so that the first and seventh loops are in close proximity with each other, thus forming a a pocket that contains the 11-cis retinal that is linked to the amino acid lysine in position 296 of the opsin molecule.
The Phototransduction Cascade is initiated when light strikes rhodopsin and isomerizes the chromophore of rhodopsin, 11-cis retinal, to all-trans-retinal to cause a conformational change in the rhodopsin molecule which results in the Photo- excited rhodopsin, Metarhodopsin II.
There are two routes that Metarhodopsin II can be directed:
In its inactive state (under dark conditions of illumination)transducin binds GDP in the form (Ta.GDP and TßY
When Metarhodopsin II reacts with transducin several reactions take place:
cGMP PDE is an enzyme consisting of four polypeptides, alpha (a, beta ßand two gamma yy polypeptides (PDEaßyy).
The reaction of Ta with PDE aßyy causes the following changes:
cGMP plays a very important role in vision. Under normal dark conditions of illumination, cGMP keeps certain channels called "cGMP-gated cationic channels", located in the outer segment cell membranes open.
In the dark, these channels allow sodium and calcium ions to enter the photoreceptor cell. Calcium ions are extruded through a calcium ion/sodium ion exchanger located in the outer segment cell membrane.
Under dark conditions of illumination, sodium enters the photoreceptor outer segment through the cGMP-gated cationic channels, travels to the inner segment where it is extruded to the outside by a sodium-potassium activated adenosine triphosphatase located in the inner segment plasma membrane.
The movement of sodium ions from the outer segment to the inner segment causes the formation a small electric current called the "dark current", between the two segments of the photoreceptor cell thus keeping the photoreceptor cell depolarized.
Exposure to light, causes the activation of the enzyme cGMP PDE which hydrlyzes cGMP thus causing a decrease in the concentration of the mononucleotide in the outer segments. This:
In order for vision to continue, the concentration of cGMP must be replenished. This is accomplished by
As mentioned above, calcium enters the outer segments under dark conditions of illumination, through the cGMP-gated cationic channels. When light strikes rhodopsin the phototransduction cascade hydrolyzes cGMP and the cGMP-gated channels are closed and the entry of calcium ions into the outer segments is blocked.
Although calcium ceases to enter the outer segments under light conditions of illumination,it continues to be extruded via a calcium ion/sodium ion exchanger located in the outer segment membrane thus reducing its concentration within the outer segments.
The decrease of calcium concentration in the outer segments, activates the enzyme, guanylate cyclase whose activity leads to the increase in the concentration of cGMP, thus opening the cGMP-gated channels and the re-entry of sodium and calcium ions. Continued esposure to light leads to the recycling of the phototransduction cascade.
Return of Rhodopsin to the Dark State
Three reactions must take place in order to return to the dark state :
As mentioned above the concentration of cGMP is increase by the activation of guanylate cyclase and the inhibition of cGMP PDE by Ta.GDP which would recombine with Tßy to react again and again with metarhodopsin II.
To return to the dark state, the ability of metarhodopsin II to combine with transducin would eventually be significantly reduced by phosphorylation with the enzyme, rhodopsin kinase followed by combination with the protein arrestin. These reaction cause the release of alltrans retinal from the protein of rhodopsin, opsin.
Absorption of light by rhodopsin in the disc membrane:
Metarhodopsin II reacts catalytically with transducin until its activity is quenched by phosphorylation by rhodopsin kinase and the binding of the phosphorylated product with the protein,arrestin.
The reactions cause the breakup of the metarhodopsin-phosphate-arrestin into:
All-trans retinal now undergoes several chemical reactions in order to be converted to 11-cis retinal and then bind to opsin and become part of the rhodopsin molecule:
I am indebted to Mrs. Helene Dussiaume, Canadian Institute for Science and Technology Information (CISTI), Ottawa, Ontario, Canada, for searching the literature and making the references available to me.
Dr. Gilles Desroches, 335 Kent St., Ottawa, Ontario, contributed valuable illustrations for this Page.
This Page is under construction and will be updated regularly. Please visit often and send me your constructive criticism: HERE