

Maintenance of diversity - coexistence in ecological communities. The diversity we observe in the natural world represents the outcome of multiple interactions between species and in relation to the abiotic environment. Ecologists have long been interested in mechanisms that allow species with similar requirements to co-occur in space and time.
I study some of the many ways in which competitive interactions can be altered to increase (or decrease) the chance that species will coexistence.
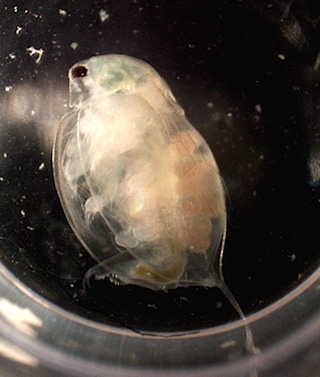
Variation between individuals of the same species, such as caused by local adaptation, phenotypic plasticity, and maternal effects, may alter the likelihood that a species can establish in a community. This has clear implications for predicting invasion, range shifts and climate change. We are using Daphnia magna and other zooplankton to explore the implications of maternal effects for coexistence and use theory, field work, and experiments to predict when such forms of non-genetic phenotypic differences may be important for community dynamics.
Other work on coexistence in ecological communities:
—With Marc Cadotte, the role of variability in fire events in maintaining Mediterranean shrub diversity and the implications of this for fire management.
—With Tadashi Fukami, using microbial microcosms, examined whether abiotic variability can alter the effects of arrival order (priority effects).
—With Peter Abrams and Ben Gilbert, explored whether the storage effect (a common variability-dependent coexistence mechanism) is expected to evolve in response to temporal variation.
Quantifying diversity. Diversity has many forms beyond species richness. Global changes in climate and habitat destruction makes conservation science a balancing act between limited resources and seemingly unlimited demand. As such, we must prioritize both species, and the underlying genetic and functional diversity they encompass.
My particular focus has been on understanding how evolutionary history is distributed through space, how evolutionary history (e.g. phylogenetic diversity) relates to taxonomic richness and thus its values for conservation prioritization, and most importantly how to correctly interpret and apply measures of evolutionary history.
Current work with sPhy, a iDiv-sponsored working group, aims to make sense of the massive number of phylogenetic metrics currently being used in ecology and conservation biology. We emphasize correct interpretation and application of metrics with reference to evolutionary history.
—In Tucker et al. (2012), my coauthors and I contest the idea that species richness is a useful proxy for phylogenetic diversity, as is sometimes contended. For example, our work shows that phylogenetic and species diversity varied spatially in the Cape Floristic Region of South Africa, and particularly varied in terms of how well different components of diversity were represented in existing reserves.
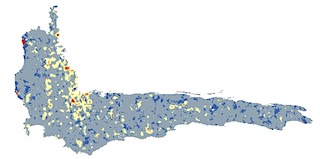
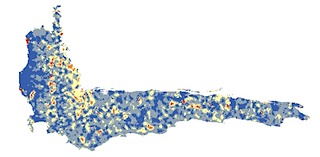
Proteaceae species richness (top) compared to phylogenetic diversity (bottom) in the Cape Floristic Region, SA.
—Further work has focused on developing a more mechanistic understanding of the relationship between species richness and phylogenetic diversity, as well as other metrics of diversity. Marc Cadotte and I develop a framework that predicts how different diversity prioritization metrics (species richness, phylogenetic diversity, and abundance-weighted versions of these metrics) are related to one another, showing that there are clear expectations based on the evolutionary history of the region, size of the regional species pool, and spatial distribution of diversity.