PSEUDOGENES
What are they, where do they come from and what can we learn from them?
![]()
PSEUDOGENESWhat are they, where do they come from and what can we learn from them?
|
|
Creation vs. Evolution:
|
What are pseudogenes? Pseudogenes are DNA sequences present in a given population, characterized by close similarities to paralogous genes and non-functionality (Mighell et al., 2000). They contain important defects that make them incapable of producing proteins in transcription and/or translation stages, or in a different scenario, they might produce a protein which is functionally different than the one produced by a normal paralog gene (Gibson, 1994). Non-functionality in pseudogenes is due to several reasons such as lacking start codons, lacking or having an extra stop codon, and altered sequence or flanking regulatory elements, all of which are necessary in normal genes for conducting proper transcription (Gibson, 1994). It is thought that mutations in pseudogenes are neutral and free from selection, and this is mainly due to the fact that they are, in most cases, located in loci where they do not cause a deleterious effect (Mighell et al., 2000).
Pseudogenes
arise by two fundamentally different ways: 1)
Retrotransposition: Retrotransposition
is one of the basic processes by which pseudogenes arise. Almost 80% of
pseudogenes have been generated in this way (Mighell et al., 2000). A brief
description of this process might be helpful to better understand how
pseudogenes emerge. The processed
mRNA of functional genes may serve as a template, from which a double
stranded DNA, called retrotransposon, is generated (Vanin, 1985).
Retrotransposons have the ability to insert themselves into the genomic DNA, a
process called retrotransposition. Although typical transposable elements carry
functional sequences for the purpose of their insertion, some retrotransposons
in humans do not have such sequences. This implies that human cells may possess an
intrinsic reverse transcription activity, providing a
necessary tool for generation of pseudogenes (Dhellin et al., 1997).
Retrotransposition
may have different outcomes, depending on the effect of natural selection
pressure on the newly arisen phenotype. It could potentially be fatal to
the entire organism, if retrotransposed sequences interrupt genes with crucial functions in life (Mighell et
al., 2000). Obviously, with the deletion of organisms carrying the defective
gene, the retrotransposed element would not be viable, either.
Retrotranspositions occurring in the non-coding regions of genomic DNA, however,
are not subjected to the immediate natural selection pressure and survive in the
form of non-functional pseudogenes. Transcription does not take place
from processed pseudogenes, since they lack the crucial 5’-promoter
sequence as well as introns, which may also contain transcription
initiation sites. Processed pseudogenes, however, contain 3’- polyadenylation
tract and flanking direct repeats, sequences presumably involved in
the process of insertion (Mighell et al., 2000). Rarely, are
retrotransposons maintained as functional, intronless genes. PGK-2
in humans and Amd-2 in mice are the two functional retrotransposed genes
corresponding to PGK-1 and Amd-1, respectively. In either case,
protein produced by the intronless retrotransposed gene is highly homologous to the one produced by the original
gene (Mighell et al., 2000) In short, a
number of pseudogenes arise by the insertion of retrotransposons in non-coding
regions of the genomic DNA. It is very unlikely that processed pseudogenes
change the organism’s phenotype, mainly because of two reasons. First they are
basically not transcribed and cannot produce proteins, and second, they result
from retrotransposition in non-coding regions; hence no pre-existing
phenotype is altered. In a few instances, however, retrotransposition gives rise
to functional intronless genes. 2)
Duplication of genomic DNA: A less common
mechanism for pseudogene generation relies on gene duplication. Gene
duplication is a source of genetic diversity which is thought to provide the
species with adaptability to the changing environment. All members of a gene
family originate from an ancestral gene. Their functionality, however, take on
different fates as mutations start to accumulate over time. Some acquire
novel adaptive functions in parallel with the environmental changes. Others may
maintain their original function, whereas the rest lose their functions and
become silent. This last group is, in fact, pseudogenes which have lost their
functions either due to conferring a disadvantageous phenotype or due to not
giving a selection advantage (Mighell et al., 2000). Hox genes belong to
a family of genes with developmental roles and are highly conserved between
different species. Two Hox genes have turned into pseudogenes during
the evolution of the puffer fish, while they are ubiquitously expressed in other
species (Meyer, 1998). Sometimes,
emergence of pseudogenes by DNA duplication does not result from accumulation of
mutations. Rather, the duplicated gene might be defective and non-functional
from
the very moment that duplication takes place. For instance, yBRCA1
(y denotes
pseudogene) arises from a tandem duplication of the 5’-end of the BRCA1 gene.
This pseudogene contains only exons 1A, 1B, and 2 of the intact gene, and
presumably, has never been functional (Brown et al., 1996). In summary, DNA
duplication serves as the second most important mechanism through which
pseudogenes evolve. DNA duplication gives rise to a series of homologous
sequences whose functions are determined by accumulation of mutations as well as
natural selection pressure from the environment. Intuitively, those sequences
with unfavorable functions are doomed to be silenced into pseudogenes or the
organisms cannot survive.
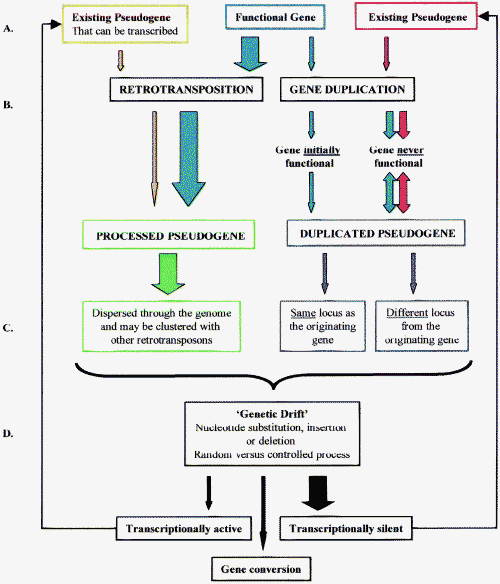
Fig 1: A schematic summary, to be interpreted in conjunction with the text, of the principal issues covered. The width of the arrows gives a rough guide to the relative importance of the different pathways at each level. A: Origin of pseudogenes. The majority of vertebrate pseudogenes are probably derived from functional genes. B: Mechanism of pseudogene formation. The majority of vertebrate pseudogenes are a result of retrotransposition of transcripts derived from genes that encode functional proteins. C: Pseudogene location. Pseudogenes persist in parts of the genome where they do not have a deleterious effect on fitness of the organism. D: Pseudogene fate. Most pseudogenes undergo genetic drift and are never transcribed. By contrast, in some instances there appears to selectional pressures that prevent major changes to the pseudogene sequence. A few pseudogenes are involved in gene conversion and a few can be transcribed. Accordingly, not all pseudogenes are unequivocally functionless.
(Re-printed with permission of FEBS Letters and Dr. A.J. Mighell).
|
|
All Rights reserved, Copyright ©2002
T0701C - JLM349S
|